Cifically cleaves unfolded protein chains. We investigated cytochrome C as a model substrate for TL’s activity and specificity for unfolded proteins. Cytochrome C can be specifically obtained in two solubleFast Proteolysis Assay FASTppFigure 2. Thermolysin is active from 206C to 806C. Thermal dependence of intrinsic rates of proteolysis of TL from 20uC to 80uC. After extensive thermal pre-equilibration, we determined kinetic traces of cleavage of an unstructured model substrate of TL, in triplicate for each temperature. The obtained rates were extrapolated based on empirically derived 4EGI-1 biological activity formulas to final concentrations of 0.1 g/L and 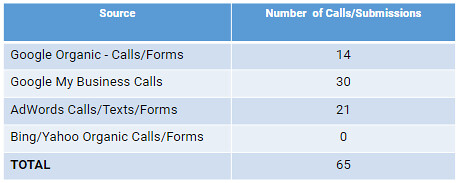 on top 0.01 g/L of TL [1]. doi:10.1371/journal.pone.0046147.gstates: either unfolded without heme or folded in the presence of heme [14,15]. We tested whether we could distinguish both forms of cytochrome C by FASTpp. TL cleaved unfolded apo cytochrome C already at 4uC whereas folded, heme-bound cytochrome C was TL-resistant up to 60uC, in agreement with earlier studies [14] (Fig. 3A, B). TL digested specifically the unfolded but not the folded protein. We concluded that TL is a KDM5A-IN-1 site suitable protease for FASTpp.Figure 3. TL preferentially cleaves unfolded protein. A, Thermal TL resistance of apo Cytochrome C (CytC). Only in absence of protease at 4uC, apo CytC was detected on the gel. Upon addition of TL, cleavage already occured on ice and no protease resistant protein was observed after additional incubation at higher temperatures. B, Thermal protease resistance of CytC in presence of the bound heme ligand. A strong band of CytC is visible in absence and presence of TL from temperatures from 4uC to 60uC. doi:10.1371/journal.pone.0046147.gKinetic protein stability can be probed by FASTpp at variable tmWe now investigated how the apparent thermal unfolding transition in FASTpp is affected by tm. For this we varied tm from 6 s to 600 s. In parallel with a step-wise increase in tm, MBP 15755315 digestion started at successively lower temperature. For instance at tm = 6 s, the unfolding occurred at 60uC while increasing tm to 600 s lowered the unfolding temperature to 49uC (Fig. 5A ). Because all assay parameters are kept constant except for tm, we can monitor kinetic stability with this assay. Proteins are “kinetically-stable” under conditions where the unfolding is slow relative to the measurement time. For instance, MBP is kineticallystable at 40uC and kinetically-unstable at 60uC for all tm values we analysed.FASTpp is insensitive to variation of TL concentrationTo refine the experimental parameters, we selected Maltose Binding Protein (MBP) as a substrate for FASTpp because it is structurally well-characterised and folds both in the presence and absence of ligand. We first probed the influence of TL concentration over four orders of magnitude on the apparent thermal melting temperature of MBP using a gradient of 50uC to 70uC and constant tm (6 s) [16]. At the lowest TL concentration of 0.001 g/L, no detectable cleavage of MBP occurred (Fig. 4A). From 0.01 to 1 g/L TL (340 nM ?4 mM), we observed loss
on top 0.01 g/L of TL [1]. doi:10.1371/journal.pone.0046147.gstates: either unfolded without heme or folded in the presence of heme [14,15]. We tested whether we could distinguish both forms of cytochrome C by FASTpp. TL cleaved unfolded apo cytochrome C already at 4uC whereas folded, heme-bound cytochrome C was TL-resistant up to 60uC, in agreement with earlier studies [14] (Fig. 3A, B). TL digested specifically the unfolded but not the folded protein. We concluded that TL is a KDM5A-IN-1 site suitable protease for FASTpp.Figure 3. TL preferentially cleaves unfolded protein. A, Thermal TL resistance of apo Cytochrome C (CytC). Only in absence of protease at 4uC, apo CytC was detected on the gel. Upon addition of TL, cleavage already occured on ice and no protease resistant protein was observed after additional incubation at higher temperatures. B, Thermal protease resistance of CytC in presence of the bound heme ligand. A strong band of CytC is visible in absence and presence of TL from temperatures from 4uC to 60uC. doi:10.1371/journal.pone.0046147.gKinetic protein stability can be probed by FASTpp at variable tmWe now investigated how the apparent thermal unfolding transition in FASTpp is affected by tm. For this we varied tm from 6 s to 600 s. In parallel with a step-wise increase in tm, MBP 15755315 digestion started at successively lower temperature. For instance at tm = 6 s, the unfolding occurred at 60uC while increasing tm to 600 s lowered the unfolding temperature to 49uC (Fig. 5A ). Because all assay parameters are kept constant except for tm, we can monitor kinetic stability with this assay. Proteins are “kinetically-stable” under conditions where the unfolding is slow relative to the measurement time. For instance, MBP is kineticallystable at 40uC and kinetically-unstable at 60uC for all tm values we analysed.FASTpp is insensitive to variation of TL concentrationTo refine the experimental parameters, we selected Maltose Binding Protein (MBP) as a substrate for FASTpp because it is structurally well-characterised and folds both in the presence and absence of ligand. We first probed the influence of TL concentration over four orders of magnitude on the apparent thermal melting temperature of MBP using a gradient of 50uC to 70uC and constant tm (6 s) [16]. At the lowest TL concentration of 0.001 g/L, no detectable cleavage of MBP occurred (Fig. 4A). From 0.01 to 1 g/L TL (340 nM ?4 mM), we observed loss  of thermal proteolysis resistance at 59uC (Fig. 4B ). Assuming comparable cleavage kinetics of the model peptide substrate and unfolded protein, we expected a minimal required cleavage time of approximately 6 s at 0.01 g/L TL to quench the unfolded fraction of protein under these conditions. Our TL titration results validated this theoretical prediction. Interestingly, at 0.01 g/ L TL, we detected unfolding and concomitant.Cifically cleaves unfolded protein chains. We investigated cytochrome C as a model substrate for TL’s activity and specificity for unfolded proteins. Cytochrome C can be specifically obtained in two solubleFast Proteolysis Assay FASTppFigure 2. Thermolysin is active from 206C to 806C. Thermal dependence of intrinsic rates of proteolysis of TL from 20uC to 80uC. After extensive thermal pre-equilibration, we determined kinetic traces of cleavage of an unstructured model substrate of TL, in triplicate for each temperature. The obtained rates were extrapolated based on empirically derived formulas to final concentrations of 0.1 g/L and on top 0.01 g/L of TL [1]. doi:10.1371/journal.pone.0046147.gstates: either unfolded without heme or folded in the presence of heme [14,15]. We tested whether we could distinguish both forms of cytochrome C by FASTpp. TL cleaved unfolded apo cytochrome C already at 4uC whereas folded, heme-bound cytochrome C was TL-resistant up to 60uC, in agreement with earlier studies [14] (Fig. 3A, B). TL digested specifically the unfolded but not the folded protein. We concluded that TL is a suitable protease for FASTpp.Figure 3. TL preferentially cleaves unfolded protein. A, Thermal TL resistance of apo Cytochrome C (CytC). Only in absence of protease at 4uC, apo CytC was detected on the gel. Upon addition of TL, cleavage already occured on ice and no protease resistant protein was observed after additional incubation at higher temperatures. B, Thermal protease resistance of CytC in presence of the bound heme ligand. A strong band of CytC is visible in absence and presence of TL from temperatures from 4uC to 60uC. doi:10.1371/journal.pone.0046147.gKinetic protein stability can be probed by FASTpp at variable tmWe now investigated how the apparent thermal unfolding transition in FASTpp is affected by tm. For this we varied tm from 6 s to 600 s. In parallel with a step-wise increase in tm, MBP 15755315 digestion started at successively lower temperature. For instance at tm = 6 s, the unfolding occurred at 60uC while increasing tm to 600 s lowered the unfolding temperature to 49uC (Fig. 5A ). Because all assay parameters are kept constant except for tm, we can monitor kinetic stability with this assay. Proteins are “kinetically-stable” under conditions where the unfolding is slow relative to the measurement time. For instance, MBP is kineticallystable at 40uC and kinetically-unstable at 60uC for all tm values we analysed.FASTpp is insensitive to variation of TL concentrationTo refine the experimental parameters, we selected Maltose Binding Protein (MBP) as a substrate for FASTpp because it is structurally well-characterised and folds both in the presence and absence of ligand. We first probed the influence of TL concentration over four orders of magnitude on the apparent thermal melting temperature of MBP using a gradient of 50uC to 70uC and constant tm (6 s) [16]. At the lowest TL concentration of 0.001 g/L, no detectable cleavage of MBP occurred (Fig. 4A). From 0.01 to 1 g/L TL (340 nM ?4 mM), we observed loss of thermal proteolysis resistance at 59uC (Fig. 4B ). Assuming comparable cleavage kinetics of the model peptide substrate and unfolded protein, we expected a minimal required cleavage time of approximately 6 s at 0.01 g/L TL to quench the unfolded fraction of protein under these conditions. Our TL titration results validated this theoretical prediction. Interestingly, at 0.01 g/ L TL, we detected unfolding and concomitant.
of thermal proteolysis resistance at 59uC (Fig. 4B ). Assuming comparable cleavage kinetics of the model peptide substrate and unfolded protein, we expected a minimal required cleavage time of approximately 6 s at 0.01 g/L TL to quench the unfolded fraction of protein under these conditions. Our TL titration results validated this theoretical prediction. Interestingly, at 0.01 g/ L TL, we detected unfolding and concomitant.Cifically cleaves unfolded protein chains. We investigated cytochrome C as a model substrate for TL’s activity and specificity for unfolded proteins. Cytochrome C can be specifically obtained in two solubleFast Proteolysis Assay FASTppFigure 2. Thermolysin is active from 206C to 806C. Thermal dependence of intrinsic rates of proteolysis of TL from 20uC to 80uC. After extensive thermal pre-equilibration, we determined kinetic traces of cleavage of an unstructured model substrate of TL, in triplicate for each temperature. The obtained rates were extrapolated based on empirically derived formulas to final concentrations of 0.1 g/L and on top 0.01 g/L of TL [1]. doi:10.1371/journal.pone.0046147.gstates: either unfolded without heme or folded in the presence of heme [14,15]. We tested whether we could distinguish both forms of cytochrome C by FASTpp. TL cleaved unfolded apo cytochrome C already at 4uC whereas folded, heme-bound cytochrome C was TL-resistant up to 60uC, in agreement with earlier studies [14] (Fig. 3A, B). TL digested specifically the unfolded but not the folded protein. We concluded that TL is a suitable protease for FASTpp.Figure 3. TL preferentially cleaves unfolded protein. A, Thermal TL resistance of apo Cytochrome C (CytC). Only in absence of protease at 4uC, apo CytC was detected on the gel. Upon addition of TL, cleavage already occured on ice and no protease resistant protein was observed after additional incubation at higher temperatures. B, Thermal protease resistance of CytC in presence of the bound heme ligand. A strong band of CytC is visible in absence and presence of TL from temperatures from 4uC to 60uC. doi:10.1371/journal.pone.0046147.gKinetic protein stability can be probed by FASTpp at variable tmWe now investigated how the apparent thermal unfolding transition in FASTpp is affected by tm. For this we varied tm from 6 s to 600 s. In parallel with a step-wise increase in tm, MBP 15755315 digestion started at successively lower temperature. For instance at tm = 6 s, the unfolding occurred at 60uC while increasing tm to 600 s lowered the unfolding temperature to 49uC (Fig. 5A ). Because all assay parameters are kept constant except for tm, we can monitor kinetic stability with this assay. Proteins are “kinetically-stable” under conditions where the unfolding is slow relative to the measurement time. For instance, MBP is kineticallystable at 40uC and kinetically-unstable at 60uC for all tm values we analysed.FASTpp is insensitive to variation of TL concentrationTo refine the experimental parameters, we selected Maltose Binding Protein (MBP) as a substrate for FASTpp because it is structurally well-characterised and folds both in the presence and absence of ligand. We first probed the influence of TL concentration over four orders of magnitude on the apparent thermal melting temperature of MBP using a gradient of 50uC to 70uC and constant tm (6 s) [16]. At the lowest TL concentration of 0.001 g/L, no detectable cleavage of MBP occurred (Fig. 4A). From 0.01 to 1 g/L TL (340 nM ?4 mM), we observed loss of thermal proteolysis resistance at 59uC (Fig. 4B ). Assuming comparable cleavage kinetics of the model peptide substrate and unfolded protein, we expected a minimal required cleavage time of approximately 6 s at 0.01 g/L TL to quench the unfolded fraction of protein under these conditions. Our TL titration results validated this theoretical prediction. Interestingly, at 0.01 g/ L TL, we detected unfolding and concomitant.
